Journal of Medical Sciences and Health
DOI: 10.46347/jmsh.v12.i2.25.407
Year: 2026, Volume: 12, Issue: 2, Pages: 159-165
Original Article
Sauliha Raq 1, Mahak Mushtaq 1, Munibul Rehman 1, Ghulam M Bhat 2
1Senior Resident, Department of Anatomy, Government Medical College, Srinagar, 190001, J&K, India.
2Professor, Department of Anatomy, Government Medical College, Srinagar, 190001, J&K, India.
Address for correspondence: Ghulam M Bhat, Professor, Department of Anatomy, Government Medical College, Srinagar, 190001, J&K, India.
E-mail: [email protected]
Received Date:09 December 2025, Accepted Date:03 May 2026, Published Date:18 May 2026
Background: Arsenic is a recognized environmental toxicant with immunotoxic and gonadotoxic potential. The spleen and testis are particularly vulnerable, and early microscopic changes may precede grossly evident pathology. The present study aimed to assess the time- and dose-dependent effects of sodium arsenate exposure on the gross and microscopic features of the spleen and testes in male albino rats.
Methods: Male albino rats were divided into control (Group A), moderate-dose (Group B), and high-dose (Group C) arsenic exposure groups. Animals were sacrificed at 4, 8, and 12 weeks for detailed gross and histopathological evaluation of spleen and testis.
Results: At 12 weeks, Group A remained normal, Group B showed follicular depletion and marginal zone changes microscopically, and Group C displayed splenomegaly, congestion, discoloration, and microscopic red pulp expansion with follicular depletion. For testes, at 4 weeks all groups retained gross and microscopic integrity. At 8 weeks, Groups A and B were unremarkable, while Group C exhibited microscopic derangement despite normal gross appearance. By 12 weeks, Group A remained unchanged, Group B showed reduced size, altered consistency, decreased Leydig cell density, interstitial widening, and seminiferous vacuolization, and Group C demonstrated gross atrophy with vascular congestion, architectural disruption, basement membrane damage, interstitial widening, Leydig cell depletion, and loss of germinal stratification.
Conclusion: Sodium arsenate induces progressive, dose- and time-dependent damage to the spleen and testes in male albino rats. Microscopic changes precede gross alterations, highlighting the need for histological assessment to detect early arsenic toxicity.
Heavy metals are naturally associated with the earth’s crust and, unlike many organic pollutants, cannot be degraded or destroyed. Their persistence, high toxicity, and bioaccumulative potential enable them to concentrate within organisms and undergo biomagnification through the food chain, making them some of the most severe biological poisons[1]. Among these metals, arsenic stands out as one of the most widespread environmental pollutants and a well-recognized human carcinogen, characterized by its complex metabolism and diverse toxic effects[2]. Arsenic contamination is a major global environmental concern due to the highly toxic and carcinogenic nature of trivalent arsenic (As³⁺, arsenate), which poses serious
implications for the environment, agriculture, and human as well as animal health[3, 4]. Human exposure primarily occurs through two major pathways: natural geological sources and anthropogenic activities[5]. Among these, atmospheric air, contaminated drinking water, and food are the principal contributors to arsenic entry into the body[6]. Industrial applications have also played a significant role, as arsenic has historically been used in wood preservatives, insecticides, fungicides, paints, herbicides, and cotton desiccants. In addition, arsenic-based compounds were once incorporated into livestock feed for pigs and poultry to enhance meat color, increase feed efficiency, and promote growth. However, due to mounting evidence of arsenic’s toxicity and associated health hazards, such practices have been increasingly restricted[7]. Arsenic exists predominantly in its inorganic forms arsenate (As³⁺) and arsenate (As⁵⁺) both of which are highly toxic to humans and terrestrial animals[8]. Among the various chemical forms of arsenic, sodium arsenate is one of the most commonly encountered inorganic salts, widely used in experimental models to study arsenic-induced toxicity. It is highly soluble, easily absorbed, and capable of inducing systemic toxic effects by generating oxidative stress, impairing cellular respiration, and altering enzymatic
activities. Prolonged exposure to sodium arsenate has been shown to cause multi-organ damage and among the organs vulnerable to arsenic exposure, the spleen and testes deserve particular attention because of their central roles in immunity and reproduction
The spleen is one of the most important lymphoid organs, functioning as a filter for blood, regulating both innate and adaptive immune responses, and maintaining immune surveillance. Its unique histoarchitecture, composed of red pulp and white pulp, makes it highly susceptible to systemic toxins and inflammatory mediators[9]. Arsenic exposure has been reported to cause significant splenic alterations, including congestion, expansion of red pulp chambers, and depletion of lymphoid follicles, which may lead to impaired immune competence, increased vulnerability to infections, and deranged hematopoietic regulation[10, 11]. The testis, on the other hand, is a vital organ for male fertility, responsible for spermatogenesis and androgen synthesis. Arsenic exposure has been strongly associated with male reproductive toxicity, disrupting germ cell proliferation and differentiation, impairing Leydig and Sertoli cell function, and inducing structural degeneration of seminiferous tubules. Histopathological changes such as vascular congestion, widening of interstitial spaces, depletion of Leydig cells, vacuolization of tubules, and reduced stratification of germinal epithelium are commonly observed[12, 13]. Such alterations not only impair spermatogenesis and reduce fertility potential but may also contribute to long-term endocrine dysfunction. Rodent models, particularly male albino rats, serve as reliable systems for assessing such toxicological effects due to their physiological and pathological resemblance to humans. Evaluating spleen and testis at sequential intervals provides critical information into the temporal progression of arsenic-induced injury. In this context, the present study aimed to observe the gross and microscopic changes in the spleen and testes of male albino rats following sodium arsenic exposure at 4, 8, and 12 weeks, thereby clarifying the dose- and time-dependent relationship between arsenic toxicity, immunological compromise, and reproductive impairment.
The present experimental study was carried out in the Postgraduate Department of Anatomy in collaboration with the Postgraduate Department of Pathology, Government Medical College, Srinagar, over a period of 18 months, of which one year was dedicated to experimental work and data collection and six months to compilation, analysis, and report writing. The study was performed on male Wistar albino rats, which were procured from the Central Animal House of Government Medical College, Srinagar. A total of eighteen healthy adult male albino rats, weighing between 180 and 200 grams, were selected for the study. Inclusion criteria comprised apparently healthy animals within the specified weight range, while those showing lethargy, poor feeding, weight loss, generalized hair loss, diarrhea, or reduced activity were excluded. Ethical approval for the use of experimental animals was obtained from the Institutional Animal Ethical Committee (IAEC/Pharma/MC/19), and all procedures were conducted in accordance with the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA).
The animals were housed in polypropylene cages of dimensions 44 × 28.6 × 30 cm³, lined with dust-free husk as bedding, and maintained under standard laboratory conditions with controlled temperature (18–29°C), humidity (30–70%), and a 12-hour light and 12-hour dark cycle. They were provided with a standard rodent diet and water ad libitum. The rats were acclimatized for one week prior to the start of the experiment, and the principles of Replacement, Reduction, and Refinement (3Rs) were strictly followed.
The rats were divided into three groups of six animals each.
Group A (Control): Received only standard diet and tap water.
Group B (Low dose): Received 50 ppm sodium arsenate (50 mg/L in drinking water) in addition to standard diet.
Group C (High dose): Received 100 ppm sodium arsenate (100 mg/L in drinking water) in addition to standard diet.
The arsenic solutions were prepared fresh each time. Animals were observed daily for any physical or behavioral changes. Sacrifices were carried out at predetermined time intervals of 4, 8, and 12 weeks under chloroform anesthesia, following CPCSEA guidelines.
After sacrifice, dissection was carried out in a designated dissection room under aseptic conditions. Gross examination included assessment of the general appearance, body weight, skin condition, physical activity, and presence of any external lesions or discharges. Following sterilization of the ventral surface with 70% ethanol, a midline incision was made from the lower jaw to the pubic symphysis to access the thoraco-abdominal cavity. Gross observations of the liver, spleen, kidneys, and testes were made, after which the spleen and testes were dissected out, rinsed in normal saline, and fixed in 10% neutral buffered formalin for histopathological evaluation.
For microscopic analysis, tissue specimens were cut into approximately 5 mm thick pieces and placed in tissue capsules labeled with organ name and group number. These were fixed in 10% formalin for 7–10 days, washed in tap water, and subjected to dehydration through ascending grades of alcohol (50%, 70%, 90%, and absolute alcohol), followed by clearing in two changes of xylene. Paraffin wax embedding was carried out at 58–60°C, with tissues left overnight for complete impregnation, and blocks were prepared using L-blocks. Sections of 6–8 μm thickness were obtained using a rotary microtome, floated in a warm water bath to remove wrinkles, and mounted on slides coated with egg albumin.
The mounted sections were stained using Harris’s Hematoxylin and Eosin. Hematoxylin imparted a blue to black coloration to nuclei, while eosin stained the cytoplasm pink and red blood cells bright red. Following dehydration through graded alcohols, clearing in xylene, and mounting in DPX, the slides were examined under a light microscope, and representative photomicrographs were taken for documentation. Gross and microscopic changes in the spleen and testes were systematically recorded for each group at 4, 8, and 12 weeks, enabling assessment of the dose- and time-dependent effects of sodium arsenate exposure on these organs.
| Time Point | Group A (Control) |
Group B (Moderate Exposure) |
Group C (High Exposure) |
|---|---|---|---|
| 4 weeks | Gross: Normal size, texture, color. Microscopy: Normal splenic architecture. |
Gross: Normal. Microscopy: Normal. |
Gross: Normal. Microscopy: Altered level of compartments. |
|
8 weeks |
Gross: Normal size, texture, color. |
Gross: Normal. |
Gross: Normal. |
|
12 weeks |
Gross: Normal size, texture, color. |
Gross: Normal. |
Gross: Enlarged, congested, discolored. |
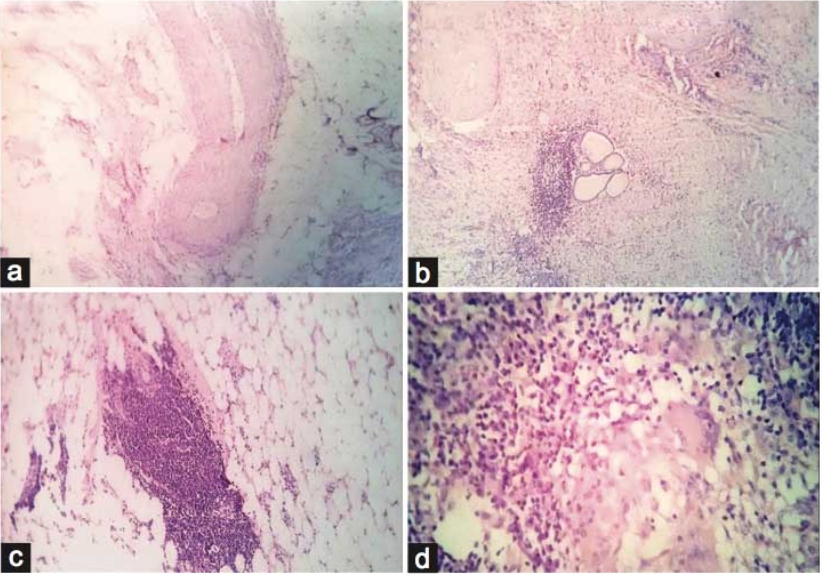
At 4 weeks, the spleen in Group A and Group B showed no gross or microscopic alterations, with preserved architecture. In Group C, the spleen appeared grossly normal, but microscopic examination revealed altered compartmental organization. At 8 weeks, the spleen in Group A remained normal in gross and microscopic appearance. Similarly, Group B did not show any significant changes. In Group C, while the gross morphology was preserved, microscopic evaluation demonstrated an increase in the red pulp chambers. At 12 weeks, the spleen in Group A continued to maintain normal size, texture, and histological integrity. In Group B, the spleen was grossly unremarkable but microscopically revealed depletion in the size and number of lymphoid follicles along with marginal zone changes. In Group C, the spleen was enlarged, congested, and discolored on gross examination, with histology showing expansion of red pulp chambers and decreased density of lymphoid follicles surrounding the central arteriole [Table. 1], [Fig. 1] & [Fig. 2].
| Time Point | Group A (Control) |
Group B (Moderate Exposure) |
Group C (High Exposure) |
|---|---|---|---|
| 4 weeks | Gross: Normal size, texture, color. Microscopy: Preserved histological architecture. |
Gross: Normal. Microscopy: Basic architecture preserved. |
Gross: Normal. Microscopy: Preserved architecture. |
|
8 weeks |
Gross: Ovoid, whitish-blue, within normal limits. |
Gross: Normal. |
Gross: Normal. |
|
12 weeks |
Gross: Normal. |
Gross: Reduced size, altered consistency. |
Gross: Atrophy observed. |
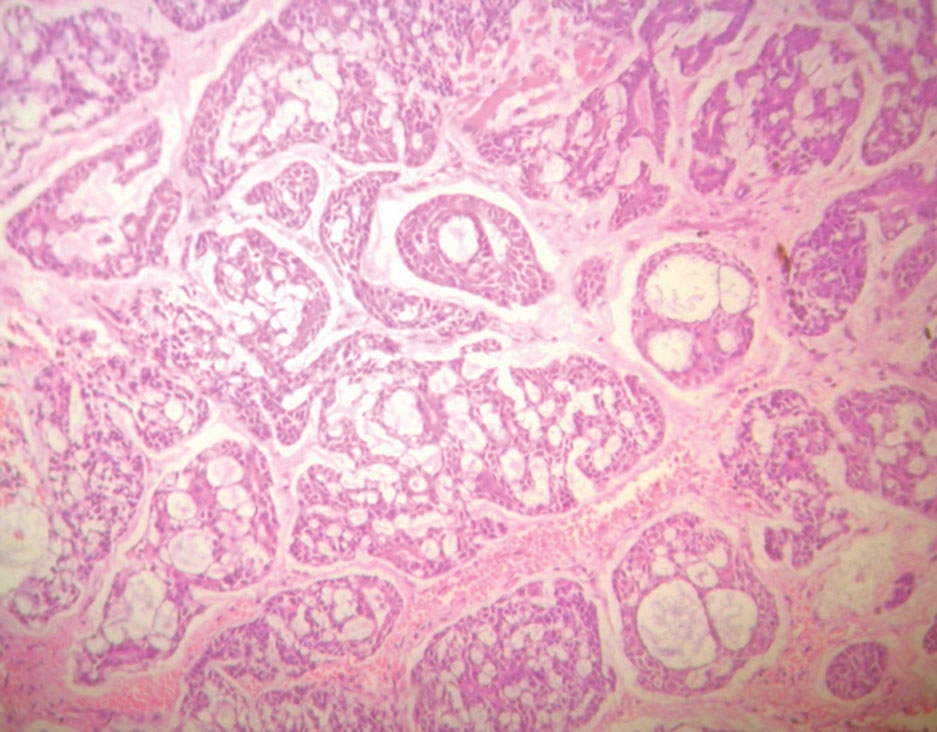
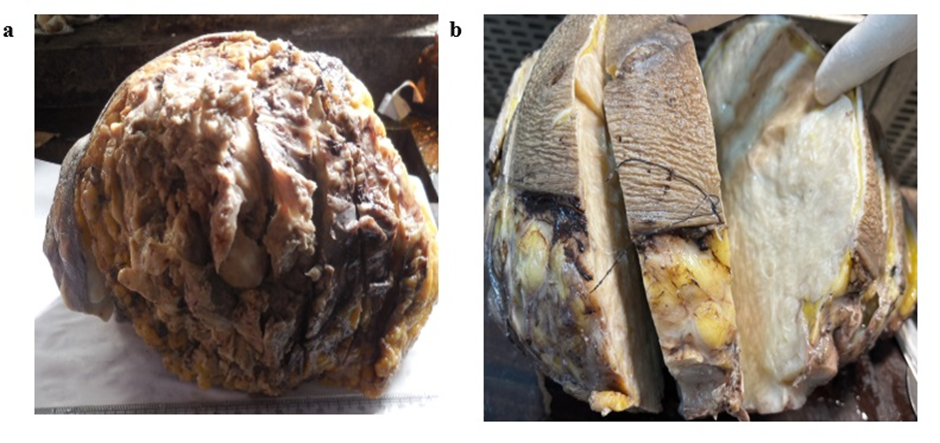
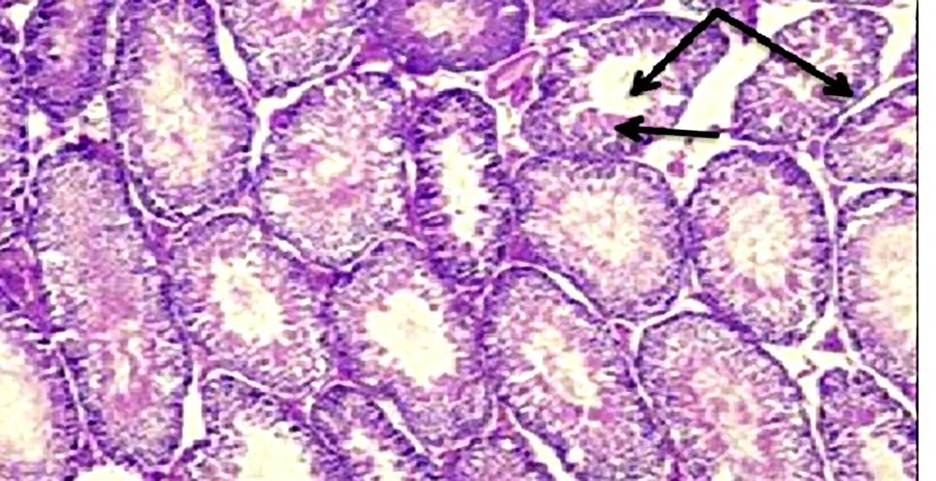
At 4 weeks, the testes in Group A and Group B appeared grossly normal with preserved histoarchitecture. In Group C, the gross features were also within normal limits, and microscopic examination demonstrated intact histological structure. At 8 weeks, the testes in Group A showed normal gross appearance and preserved histology. In Group B, the gross size, weight, and color remained normal, and the basic architecture was retained. In Group C, while the testes appeared grossly normal, histological analysis revealed deranged architecture. At 12 weeks, the testes in Group A did not exhibit any significant gross or microscopic alterations. In Group B, a reduction in testicular size and altered consistency were noted grossly, while histology showed decreased Leydig cell density, widening of interstitial spaces, and vacuolization of seminiferous tubules. In Group C, the testes exhibited gross atrophy, and microscopic examination revealed vascular congestion, architectural disruption, basement membrane damage, widening of interstitial spaces, depletion of Leydig cells, and reduced stratification of germinal epithelium [Table. 2], [Fig. 3] & [Fig. 4].
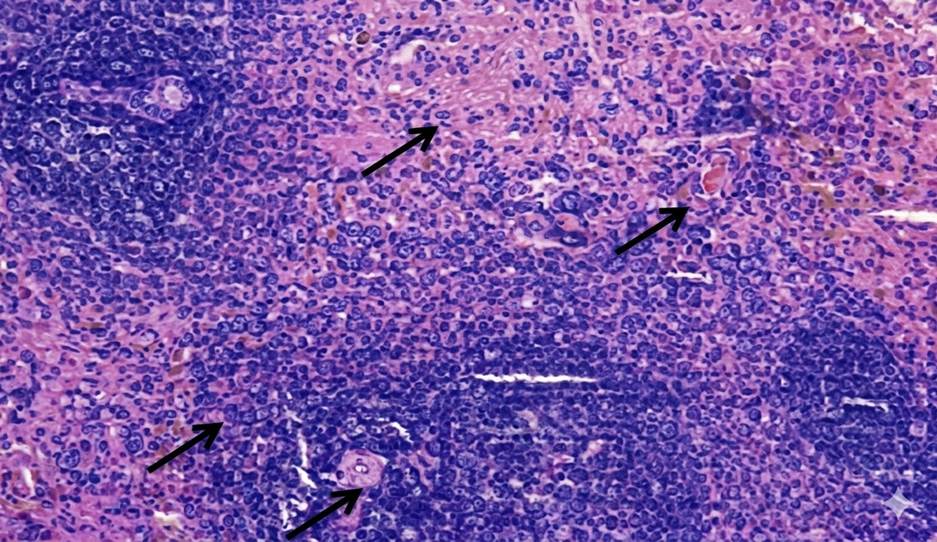
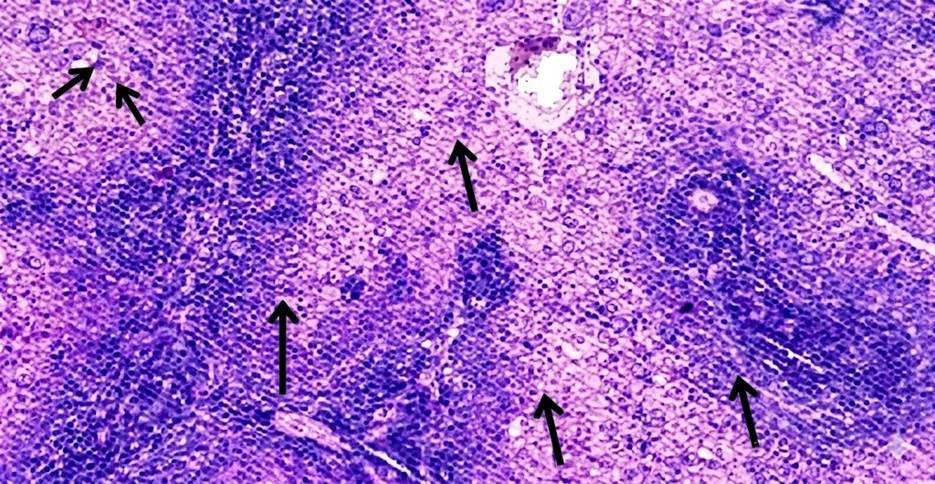
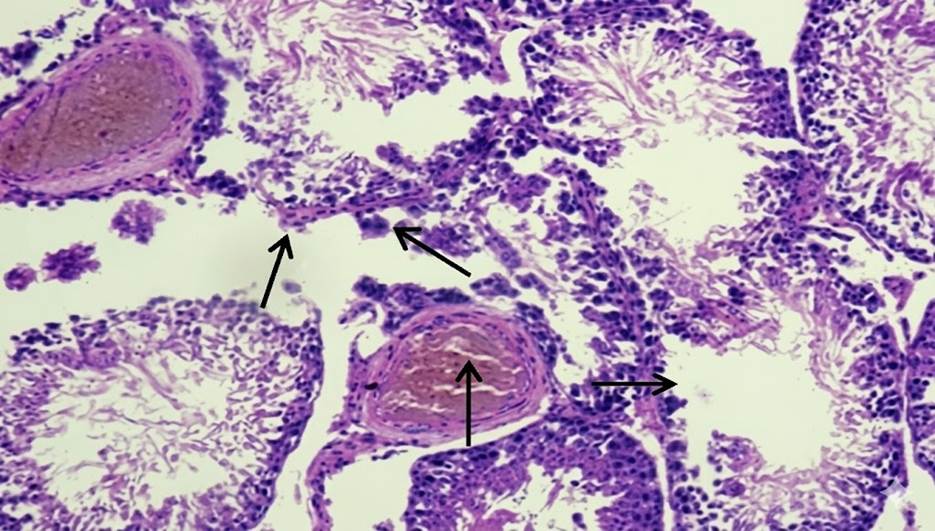
The present study demonstrated a clear time- and dose-dependent pattern of arsenic-induced splenic alterations in male albino rats. At 4 weeks, Group A and Group B showed preserved gross and microscopic architecture, whereas Group C already exhibited microscopic disorganization of splenic compartments despite a grossly normal appearance. By 8 weeks, Group C displayed expansion of red pulp chambers on histology, while the gross morphology remained unchanged. At 12 weeks, Group B revealed depletion in the size and number of lymphoid follicles along with marginal zone changes, whereas Group C exhibited gross enlargement, congestion, and discoloration, with histological evidence of red pulp expansion and reduced density of lymphoid follicles surrounding the central arteriole. These observations highlight that arsenic toxicity in the spleen progresses in a sequential manner, where microscopic changes precede gross alterations, and severity increases with both dose and duration. Our findings are consistent with the work of Yan et al., who reported that chronic arsenic exposure induces a time-dependent modulation of splenic inflammation and immunosuppression, characterized by early inflammatory changes followed by histological disorganization and functional decline[14]. Similarly, Barai et al., demonstrated splenic enlargement and red pulp disruption in mice exposed to arsenic, with the changes becoming more pronounced with prolonged exposure[15]. In agreement with our findings, Soria et al., also observed that arsenic bioaccumulation in the spleen leads to enzymatic disturbances, oxidative stress, and depletion of lymphoid elements, thereby explaining the follicular atrophy and marginal zone alterations evident in our Group B and Group C rats at later time points[16]. The expansion of red pulp and depletion of lymphoid follicles seen in our high-dose group at 12 weeks parallels the histopathological findings described by Al-Forkan M et al. and Burchiel et al., who emphasized that arsenic exposure adversely affects splenic immune parameters, including lymphoid tissue integrity and extramedullary hematopoiesis[17, 18]. Mechanistically, oxidative stress and apoptosis of splenocytes have been suggested as the primary drivers of arsenic-induced injury. In this regard, Yan et al. (2020), Zhao et al. and Soria et al., highlighted that chronic arsenic exposure disrupts redox balance and triggers lymphocyte apoptosis, leading to progressive immune dysfunction and structural disintegration of splenic tissue[14, 15, 19]. Furthermore, the dose-dependent nature of our findings, with milder alterations in Group B compared to the severe changes in Group C, aligns with the observations of Yokohira et al. (2008), who reported that both the severity and onset of arsenic-induced tissue injury are closely related to the administered dose[20]. This dose-time interaction explains why follicular depletion was confined to Group B only at the 12-week interval, while Group C manifested gross splenomegaly and severe microscopic derangements. In nutshell, our study corroborates existing literature that arsenic exposure produces progressive, dose-dependent splenic toxicity. The sequential pattern observed ranging from early compartmental disorganization to follicular depletion and red pulp expansion suggests that the spleen is highly sensitive to arsenic-induced oxidative and immunotoxic stress. These findings also emphasize the importance of microscopic evaluation for detecting early changes, which may precede overt gross abnormalities.
In the present study, the testes showed no gross or microscopic abnormality at 4 weeks across all groups. By 8 weeks, the high-dose group (Group C) already exhibited microscopic derangement despite preserved gross appearance, and by 12 weeks both moderate (Group B) and high (Group C) exposure produced progressive pathology, reduced testicular size and altered consistency in Group B and frank atrophy with extensive histological damage in Group C. This time- and dose-dependent progression, in which early microscopic changes precede later gross atrophy, has been consistently described in experimental models of inorganic arsenic toxicity. Jana et al. observed that arsenic exposure in adult rats led to germ-cell degeneration and impaired androgen production, emphasizing that microscopic tubular injury can precede gross testicular shrinkage[21]. Similarly, Kaushal et al., in their study on postnatal sodium arsenate exposure, reported vacuolization of seminiferous tubules, perturbation of Sertoli and Leydig cells, and impaired spermatogenesis, findings closely comparable to the vacuolization, interstitial widening, and loss of germinal stratification documented in the present 12-week high-dose group[22]. Histopathological hallmarks recorded in our 12-week specimens, vacuolization of seminiferous epithelium, widening of interstitial spaces, depletion of Leydig cells, basement membrane disruption, and loss of orderly germinal stratification have been repeatedly described across sodium-arsenite studies. For instance, Ahmad I et al., reported a significant decrease in seminiferous tubular diameter, thickening of basement membrane, early arrest of spermatogenesis, damaged Leydig cells, and overall testicular atrophy following high-dose arsenic exposurer, while Pachnanda et al., documented dose-dependent destruction of testicular histoarchitecture, including complete arrest of spermatogenesis, disfigured seminiferous tubules, absence of spermatids in tubular lumens, distortion of germinal epithelium, interstitial edema, and damage to Sertoli and Leydig cells, which is akin to our study[23, 24]. These observations reinforce the reproducibility of testicular lesions in arsenic toxicity models. Mechanistic studies provide information into these pathological alterations. Aitken R et al. demonstrated that arsenic induces oxidative stress in testicular tissue, depleting antioxidants and increasing lipid peroxidation, which in turn triggers mitochondrial dysfunction and apoptosis in germ cells[25]. Additional work by Yilmaz B et al. showed that arsenic disrupts Leydig cell function and reduces testosterone synthesis, while exposing testicular tissue to apoptotic and degenerative changes[26]. These findings explain the Leydig cell depletion, interstitial widening, and architectural collapse observed in our 12-week moderate- and high-dose groups. Furthermore, Sun H et al., reviewed arsenic’s interference with the hypothalamic pituitary gonadal axis, adding another mechanistic layer to explain the reproductive decline[27]. The temporal dissociation between early microscopic injury and later gross atrophy observed in this study is supported by previous work. For example, Singh et al. reported that histological abnormalities such as germinal epithelial vacuolization and spermatogenic arrest appear before significant reductions in testicular weight or gross atrophy become apparent[28]. This observation highlights that reliance solely on gross morphology may underestimate the extent of early arsenic-induced reproductive toxicity.
The observed splenic/ testicular alterations can be reasonably attributed to sodium arsenate exposure, as other potential contributory factors were carefully minimized. All animals were age-matched, maintained under identical housing, dietary, and environmental conditions, and were free from clinical illness or stress-related exclusion criteria. The absence of comparable splenic/ testicular changes in the control group across all time points further excludes handling stress, nutritional deficiency, infection, or procedural artifacts as causes, thereby strengthening the causal association between arsenic exposure and the documented splenic histopathological changes.
The present study demonstrated that arsenic exposure produces progressive alterations in both the spleen and testes of male albino rats, with microscopic changes often preceding gross abnormalities. In the spleen, early exposure was associated with subtle histological changes, while prolonged exposure led to congestion, discoloration, and architectural disruption, suggesting impaired immune function. Similarly, the testes exhibited a sequence of changes beginning with preserved gross morphology but early microscopic vacuolization and disorganization of seminiferous tubules. With extended exposure, these changes progressed to reduced testicular size, loss of Leydig cells, depletion of germinal epithelium, vascular congestion, and gross atrophy. The findings indicated that arsenic has both immunotoxic and gonadotoxic effects, which intensify with longer durations of exposure. Importantly, the study underscored that microscopic alterations appear much earlier than visible gross changes, making histopathological evaluation critical for detecting early organ damage induced by arsenic toxicity.
1. Effect of cadmium on glycogen content in muscle, liver, gill and kidney tissues of freshwater fish Channa punctatus (Bloch). Journal of Applied and Natural Science. 2019; 11 (2). Available from: https://doi.org/10.31018/jans.v11i2.2115
2. Roy P, Saha A. Metabolism and toxicity of arsenic: a human carcinogen. Current Science. 2002;82(1):38-45.
3. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. International Journal of Environmental Research and Public Health. 2018; 15 (1). Available from: https://doi.org/10.3390/ijerph15010059
4. Quercetin mediated attenuation of cadmium‐induced oxidative toxicity and apoptosis of spermatogenic cells in caprine testes in vitro. Environmental and Molecular Mutagenesis. 2021; 62 (6). Available from: https://doi.org/10.1002/em.22450
5. Arsenic-Induced Carcinogenesis: The Impact of miRNA Dysregulation. Toxicological Sciences. 2018; 165 (2). Available from: https://doi.org/10.1093/toxsci/kfy128
6. Seven potential sources of arsenic pollution in Latin America and their environmental and health impacts. Science of The Total Environment. 2021; 780 Available from: https://doi.org/10.1016/j.scitotenv.2021.146273
7. Acute and chronic arsenic toxicity. Postgraduate Medical Journal. 2003; 79 (933). Available from: https://doi.org/10.1136/pmj.79.933.391
8. Health Effects of Chronic Arsenic Exposure. Journal of Preventive Medicine and Public Health. 2014; 47 (5). Available from: https://doi.org/10.3961/jpmph.14.035
9. Structure and function of the immune system in the spleen. Science Immunology. 2019; 4 (33). Available from: https://doi.org/10.1126/sciimmunol.aau6085
10. A study on behavioral, neurotoxicological, and immunotoxicological effects of subchronic arsenic treatment in rats. Journal of Toxicology and Environmental Health, Part A. 2002; 65 (16). Available from: https://doi.org/10.1080/152873902760125390
11. Splenic cell targets in gallium arsenide-induced suppression of the primary antibody response. Toxicology and Applied Pharmacology. 1991; 110 (1). Available from: https://doi.org/10.1016/0041-008x(91)90296-q
12. Reproductive toxicity in male rats induced by chronic arsenic exposure involves hormonal and structural changes. Scientific Reports. 2025; 15 (1). Available from: https://doi.org/10.1038/s41598-025-14929-6
13. Arsenic Toxicity in Male Reproduction and Development. Development & Reproduction. 2015; 19 (4). Available from: https://doi.org/10.12717/dr.2015.19.4.167
14. Chronic arsenic exposure induces the time-dependent modulation of inflammation and immunosuppression in spleen. Cell & Bioscience. 2020; 10 (1). Available from: https://doi.org/10.1186/s13578-020-00448-6
15. Amelioration of arsenic-induced toxic effects in mice by dietary supplementation of Syzygium cumini leaf extract. Nagoya Journal of Medical Science. 2017; 79 (2). Available from: https://doi.org/10.18999/nagjms.79.2.167
16. Immunotoxicological effects of arsenic bioaccumulation on spatial metallomics and cellular enzyme response in the spleen of male Wistar rats after oral intake. Toxicology Letters. 2017; 266 Available from: https://doi.org/10.1016/j.toxlet.2016.12.014
17. A Sub-Chronic Exposure Study of Arsenic on Hematological Parameters, Liver Enzyme Activities, Histological Studies and Accumulation Pattern of Arsenic in Organs of Wistar Albino Rats. Journal of Cytology & Histology. 2016; 07 (02). Available from: https://doi.org/10.4172/2157-7099.1000s5:006
18. Immunotoxicity and biodistribution analysis of arsenic trioxide in C57Bl/6 mice following a 2-week inhalation exposure. Toxicology and Applied Pharmacology. 2009; 241 (3). Available from: https://doi.org/10.1016/j.taap.2009.09.019
19. Chronic arsenic exposure in drinking water interferes with the balances of T lymphocyte subpopulations as well as stimulates the functions of dendritic cells in vivo. International Immunopharmacology. 2019; 71 Available from: https://doi.org/10.1016/j.intimp.2019.03.022
20. Effect of Sodium Arsenite Dose Administered in the Drinking Water on the Urinary Bladder Epithelium of Female Arsenic (+3 Oxidation State) Methyltransferase Knockout Mice. Toxicological Sciences. 2011; 121 (2). Available from: https://doi.org/10.1093/toxsci/kfr051
21. Effects of chronic exposure to sodium arsenite on hypothalamo-pituitary-testicular activities in adult rats: possible an estrogenic mode of action. Reproductive Biology and Endocrinology. 2006; 4 (1). Available from: https://doi.org/10.1186/1477-7827-4-9
22. Postnatal exposure to sodium arsenite (NaAsO2) induces long lasting effects in rat testes. Toxicology International. 2012; 19 (2). Available from: https://doi.org/10.4103/0971-6580.97225
23. Arsenic-induced microscopic changes in rat testis. The Professional Medical Journal. 2008; 15 (02). Available from: https://doi.org/10.29309/tpmj/2008.15.02.2760
24. Histopathological alterations in testicular tissue of male rats exposed to arsenic. Journal of Applied and Natural Science. 2012; 4 (2). Available from: https://doi.org/10.31018/jans.v4i2.258
25. Antioxidant Systems and Oxidative Stress in the Testes. Oxidative Medicine and Cellular Longevity. 2008; 1 (1). Available from: https://doi.org/10.4161/oxim.1.1.6843
26. Disruption of Leydig cell steroidogenic function by sodium arsenite and/or sodium fluoride. Theriogenology. 2022; 193 Available from: https://doi.org/10.1016/j.theriogenology.2022.09.019
27. Mechanisms of arsenic disruption on gonadal, adrenal and thyroid endocrine systems in humans: A review. Environment International. 2016; 95 Available from: https://doi.org/10.1016/j.envint.2016.07.020
28. Singh PK, Singh J, Gupta AK, Singh AK. Effects of ground water arsenic on the testes of albino rats. Journal of Ecophysiology and Occupational Health. 2011;11(1-2):33-7.
Subscribe now for latest articles and news.