Journal of Medical Sciences and Health
Year: 2022, Volume: 8, Issue: 1, Pages: 78-87
Original Article
Vasudha Kulkarni1, H L Tejaswi2, Ashwini C Appaji3
1Associate Professor, Dr. B. R. Ambedkar Medical College, Kadugondanahalli, 560045, Bengaluru,
2Associate Professor, Adichunchanagiri Institute of Medical Sciences, B G Nagara, Bellur, 571432, Karnataka,
3Associate Professor, M S Ramaiah Medical College, Mathikere, Bengaluru, 560054
Address for correspondence: Vasudha Kulkarni, Associate Professor, Dr. B. R. Ambedkar Medical College, Kadugondanahalli, 560045, Bengaluru. E-mail: [email protected]
The human lung is a dynamic organ of constant biotransformation. Out of 40 different cell types of lungs, Type‑1 and Type‑2 pneumocytes, pulmonary capillary endothelium, and cells of lung interstitium play a vital role in functioning of the lung. Exploring the structure of these cells would shed light on the pathogenesis of various diseases affecting the lungs. A systematic literature search was conducted using three research databases – Medline, Google Scholar, and Cochrane. Out of ninety articles published between the year 1981 and 2020, 41 articles were included in the study based on the inclusion and exclusion criteria. Among the cellular components of the alveolar cell complex of the lung, the Type‑2 pneumocytes show versatility in its function in the form of surfactant synthesis, immune reaction, and regenerative capacity. The pathogenesis of the lung injury is due to the loss of functions of the Type‑2 pneumocytes and the intercellular junction functional discrepancy. Irrespective of the cause of lung disease, there is an increase in permeability in the alveolocapillary membrane leading to interstitial edema. This, in turn, leads to hyperventilation, low oxygen saturation, and a decrease in lung capacity. The pericytes present in the interstitium have a preventive role to play in lung injury and potential stem cell function.
Keywords: alveolar epithelial cells, capillary endothelium, lung interstitium, type-1 pneumocytes, type-2 pneumocytes
The human lung is not merely an organ exchanging gases but a highly metabolically active organ with more than 40 different cell types. The morphology, lung volume, and lung capacities vary with sex and ethnicity.[1] The alveolus has a surface area of 140 m2 and of 250 µm diameter.[2] An estimated 25 million alveoli at birth increase to 300 million by eight years of age.[3] Alveoli are lined by 19 × 109 cells with a predominance of Type-1 and Type2 pneumocytes.[4] Type-1 pneumocytes form the blood–air barrier, whereas Type-2 pneumocytes pro duce surfactants.[5,6] Another major stakeholder is the pulmonary endothelium.[7] The integrity of Type1 pneumocytes, Type-2 pneumocytes, pulmonary capillary endothelium, alveolar macrophages, and lung interstitium cells determines the alveoli’s inflammatory responses. Thus, we propose the term “Alveolar cell complex of the human lung” for a group of cells in and around the alveoli. The objectives of this review were to characterize the ultrastructure of Type-1 and Type-2 pneumocyte and pulmonary endothelium and establish the association between cells of the alveolar cell complex.
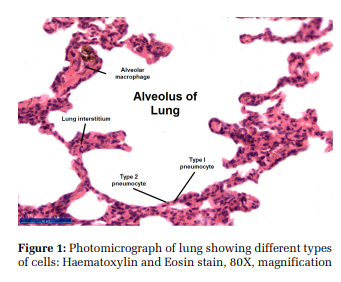
The alveolus is a mosaic of Type-1 and Type-2 pneumocytes, capillary endothelial cells, interstitial cells, and alveolar macrophages [Figure 1].[8] The interaction between these cell types triggers an inflammatory chain in the lung.
Type 1 Pneumocyte
These are simple squamous cells with thin cytoplasmic extensions spanning a surface area of 93% to 97% of the alveolus. They are 50 µm in diameter and 0.1- 0.2 µm thin.[9,10] They constitute 35%-40% of the alveolar cell population. They contain microvilli, mitochondria, rough and smooth endoplasmic reticulum.[11,12] The cytoplasm extends around the nucleus as a leaflet with terminal bars at the place of meeting of cytoplasmic leaflets.[8]
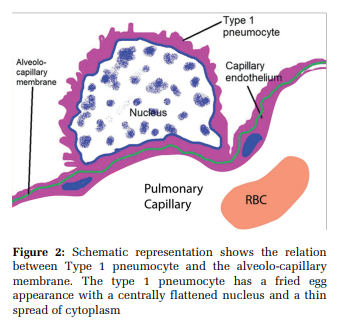
[Figure 2] The mean thickness of the blood air barrier between type-1 pneumocyte and capillary endothelium is 0.6 µm[10,13,14] with 50–100 m2 of surface area.[15] These cells contain pinocytic vesicles that help remove particles from the surface of the alveolus.[6] Aquaporin- 5 and Caveolin-1 are transmembrane protein markers specific for type-1 pneumocytes [Table 4]. Aquaporin-5 is a specific membrane-bound protein channel facilitating the exchange of gases between type-1 pneumocytes and capillary endothelium and fluid transport across alveoli.[16] Aquaporin-5 has been implicated in lung carcinogenesis.[17]
. Specific gene expressions include fibroblast growth factor receptor activating protein-1, purinergic receptor P2X7, interferoninduced protein, and Bcl2-associated protein-1.[6]
Type-2 Pneumocytes / Alveolar type-2 epithelial
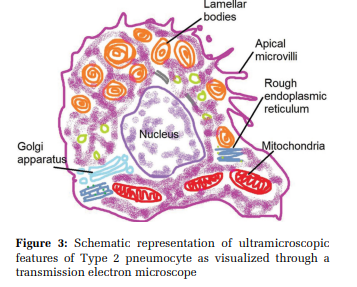
cells / Granular pneumocytes / Giant corner cells They constitute 60 % of the alveolar epithelial population and 15% of the lung parenchymal population.[16,17] They are cuboidal cells, 15 µm in diameter, containing a basal nucleus with the prominent nucleolus, lamellar bodies, multivesicular bodies, mitochondria, endoplasmic reticulum, Golgi apparatus, and apical microvilli [Figure 3].[16] Each type-2 pneumocyte contains 200- 500 lamellar bodies (1 µmin diameter) that store and release surfactant.[18,19]
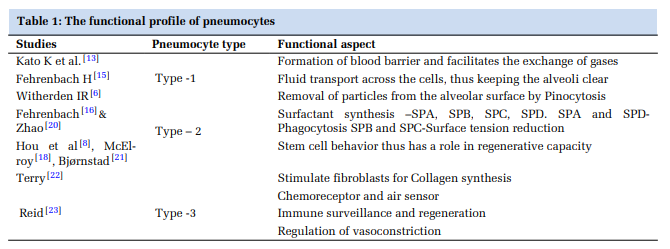
Functions of Type 2 Pneumocytes: [Table 1]
Surfactant synthesis: Surfactant is produced by type2 pneumocytes and Clara cells of bronchi. It consists of 90% phospholipids and 10% proteins. SP-B is responsible for the surface tension of alveoli at birth.[24,25] SP-C is exclusively secreted by type2 pneumocytes. Chromosome-10 has the gene for SP-A and SP-D, chromosome-2 for SP– B, and chromosome-8 for SP-C.[26] Long-term exposure to toxicants like cigarette smoking, ozone, nitrogen dioxide, sulfur dioxide, hydrogen sulfide, and dust particles affects the surfactant system. SP-A levels are significantly low in cigarette smokers.[20]
Phagocytosis: Mason, 1977 named type-2 pneumocyte as “Defender of Alveolus” as it prevents infection by modulation of alveolar hypo phase and halting transudation of interstitial fluid.[16]
Amiloride-sensitive epithelial Na channel (ENaC), Na, K-ATPase, and cystic fibrosis transmembrane regulator (CFTR) are specific ion channels of Type-2 pneumocytes helping in Na+ and Cl- transport.[11,27]
Regenerating capacity: Type-2 pneumocytes are progenitors of type-1 pneumocytes,[9] with the transformation taking 2-5 days.[8] Mice genetic lineage tracing studies have demonstrated that subgroups of type-2 pneumocytes generate alveolospheres, self-generating lung organoids.[21] High levels of telomerase activity in type-2 pneumocytes also substantiate that they have regenerative capacity.[28] Intercellular and extracellular factors affecting
pneumocyte type 2 In-Vitro cultures of type-2 pneumocytes indicated that their proliferation depended on the presence of alveolar macrophages, spindle cells, and extracellular matrix membrane.[29] The growth of type2 pneumocytes did not happen in the in-vitro cultures, which used plastic as a medium[30] due to the lack of chemical interaction between cellular and extracellular components.[4] Thus, disruption in one of the cellular components might decrease the functionality of the other cells.[31] The type-2 pneumocytes of an adult alveolus can synthesize fibronectin, antiprotease 1-antitrypsin, and type IV collagen, whereas fetal-type cells produce proteoglycans.[22,32]
Effect due to Lung Injury
In the context of lung injury, studies have shown that type 2 pneumocytes become hyperplastic. They produce tumor necrosis factor- α, IL 1, IL 6, and IL 8 (Interleukins), Tumour growth factor - β1, and platelet-derived growth factor (PDGF).[24] . These profibrotic factors produced by the hyperplastic type 2 pneumocytes induce pulmonary fibrosis, which is detrimental to the lungs.[30]
Communication between the cellular and extracellular components of the lung: Intercellular Cell junctions The adjacent type 1 pneumocytes are connected by desmosomes and tight junctions that prevent interstitial tissue fluid leakage into the alveolus.[10] The type 1 pneumocyte and type 2 pneumocyte have tight junctions (Zonula occludens). These epithelial cells determine the permeability of water-soluble particles across the alveolar-capillary membrane.[19] On the contrary, the pulmonary endothelial cells are held together by loose junctions aiding the free flow of fluid and macromolecules.[17]
Pores of Kohn and Channels of Lambert The type-2 pneumocytes lining the adjacent alveoli communicate through the alveolar duct and pore of Kohn.[16] The alveolar pores of Kohn are 3 to 13 µm in diameter. Normally, they are closed by surfactants, but macrophages can pass through them. When they enlarge in size (> 15 µm in diameter), they are called fenestrae.[10] The alveoli communicate with respiratory bronchioles through bronchoalveolar channels of Lambert (120 µm in diameter). The pores of Kohn appear by four years of birth, predominantly distributed in peribronchial, perivascular, and subpleural alveoli of apical portions of upper and lower lobes. Pores of Kohn and bronchoalveolar channels of lambert are beneficial for collateral ventilation (bypass of normal airway circulation) in chronic obstructive pulmonary disease (COPD).[22]
Cell markers of type 2 pneumocyte
Type -2 pneumocytes exhibit a spectrum of cell markers [Table 4]. Surface markers specific for type -2 pneumocytes are Maclura pomifera (MPA) and N-acetyl galactosamine. Glycoprotein (GP300) and Ca1 derived from the human laryngeal carcinoma cell line are monoclonal antibodies against surface proteins of type-2 pneumocytes. GP 300 is an autoantigen causing Heymann nephritis because it is a common marker for apical plasma membranes of renal, type-2 pneumocytes, intestinal, epidydimal, and yolk sac epithelia. Enzymes specific for type2 pneumocytes include α-galactosidase and alkaline phosphatase.[6,21,24,28]
Immunohistochemistry studies have shown that type- 2 pneumocyte markers also include MUC1 (Mucin 1- expressed by normal and neoplastic type - 2 cells), TTF1(Thyroid transcription factor 1), CD44v6 (cell surface adhesion molecule with cellular and cell-matrix interactions), and TF (Thomsen - Friedenreich antigen). These markers are diagnostic of the source of neoplastic cells in pulmonary sclerosing haemangioma. CD44v6 is expressed in saccular and canalicular lung development stages and plays a significant role in tumorigenesis.[25] Recent studies of SARS - CoV (COVID 19), reveal that type-1 and 2 pneumocytes express ACE 2 receptors on their surface. Type2 pneumocytes cell line A549 have expressed ACE 2 receptors abundantly, paving the way for Coronavirus binding, entry and replication and further damage to the alveoli.[32]
Cellular interactions of type 2 pneumocyte
SP-A from type-2 pneumocytes modulates the release of oxygen radicals and nitric oxide from macrophages. SP - A has an autocrine effect on type2 pneumocytes. Type-2 pneumocytes bear major histocompatibility complex II receptors inducing differentiation of basophils, eosinophils, neutrophils, granulocytes, and T lymphocytes.[16] These cells act as transducers of an inflammatory signal from the alveolus to the capillary bed to recruit granulocytes at the site of inflammation. Hepatocyte growth factor and heparin-binding epidermal growth factor-like protein produced by alveolar macrophages act as mitogens for type- 2 pneumocytes.[29]
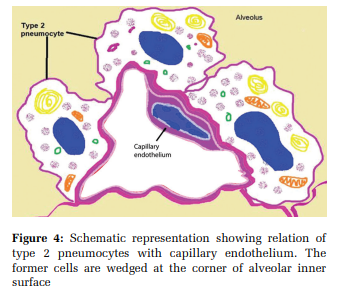
The insulin-like growth factor of type- 2 pneumocytes stimulate collagen synthesis in the fibroblasts. The surfactant lipids can influence decreased production of type- 1 collagen synthesis and trigger fibroblast apoptosis. Pulmonary endothelial cells cause a paracrine effect on surfactant production. The cellular integrity of type- 2 pneumocytes is essential for adequate functioning of pulmonary capillary endothelium [Figure 4]. Type- 2 pneumocytes produce endothelin and stimulate the production of prostaglandin E2.[30] Loss of type- 1 pneumocyte in lung alveolus triggers the production of growth factors from type- 2 pneumocytes.[10]
Type 3 Pneumocyte
It is a columnar / flask-shaped / pear-shaped cell with a broad base and apical microvillus (5-10% of alveolar cells). Each cell has 120- 140 microvilli, measuring 0.5–1 nm in length and 150–180 nm in width. The cytoplasm contains glycogen, vesicles, smooth endoplasmic reticulum at its supranuclear region, giving it a striped snake appearance. The cell is innervated by an unmyelinated nerve at its base and acts as an alveolar fluid sensor, air sensor, and chemoreceptor. Its microvilli markers are cytokeratin 18, villin, and fimbrin. The cells are concentrated at the first bifurcation of the alveolar duct. It helps in immune surveillance, cell regeneration, vasoconstriction regulation, and perfusion during hypoxia.[23]
Capillary endothelial cells
The pulmonary veins originate from the alveolar plexus and are a critical regulator of vascular homeostasis[33,34] and a major component of the alveolo-capillary unit.[35]140 m2 of the alveolar surface is covered by the pulmonary capillary, accounting for 85 % to 95% of the alveolar surface.[10] The alveolocapillary membrane is 0.1 - 0.5 µm in thickness.[36]
Ultrastructure of Pulmonary endothelium
The endothelial cell of the pulmonary artery is elliptical, whereas the endothelial cell of the pulmonary vein is polygonal. The pulmonary endothelium has apical rod-shaped microvilli.[10] The oval nuclei of endothelium lie parallel to the blood flow path.[37] They have mitochondria concentrated towards capillary branch points. The mitochondria trigger the nuclear factor-kB (Nf - kB) pathway, causing microvascular infiltration of antigens.[38] Coagulation factor VIII (Von Willebrand factor) and P - selectin are stored in cylindrical, membranebound structures called Weibel - Palade bodies. Such secretory bodies are absent in endothelial cells of pulmonary capillaries.[34] The Golgi bodies occupy < 0.5% of endothelial cell volume.[35] [Figure 4]
The total pulmonary capillary surface area is 126- 130 m2, endothelium contributing to 1100 µm2 per cell. The cytoplasm has abundant caveolae. The zone of approximation of two plasma membranes of capillary endothelium devoid of caveolae is called the avesicular zone of capillary endothelium. This zone has a thickness of 20 -30 nm.[8]
Cell junctions of Pulmonary endothelium
These junctions are occludins, claudins, focal adhesion plaques, junction adhesion molecules, cytoplasmic proteins, cytoskeletal microtubules, and actin microfilaments.[17] They function as watchmen regulating the movement of trans endothelial fluids, cells, and proteins. They determine leucocyte and macromolecular transmigration and edema formation.[10,39]
The intima of the pulmonary endothelium is coated with negatively charged glycocalyx made up of glycosaminoglycans, proteoglycans, and glycoproteins.[10] Glycocalyx coat is the gatekeeper regulating migration/ adhesion of leukocytes and inhibiting intravascular thrombosis. The coat occupies 3.7+ 0.6 % of the lumen of pulmonary capillaries. The coat can be visualized in Transmission Electron Microscopy (TEM) after staining with lanthanum nitrate.[39] Electron microscopic studies on lab mice lungs have indicated that the glycocalyx layer gets disrupted on injection of lipopolysaccharides. The loss of glycocalyx is induced by the neutrophils leading to ARDS-like symptoms and replenished with a resolution of injury/infection.[39]
Functions of pulmonary endothelium
It produces vasodilator factors such as Nitric oxide (NO), Prostaglandins (PGI2), Tranexemic acid (Tx A2), vasoconstrictors such as Endothelin1 (ET1) and serotonin (5- HT), vascular growth factors such as vascular endothelial growth factor (VEGF). High or low-pressure flow decreases NOS expression causing vasoconstriction/vasodilation.[37,40] The uniqueness of pulmonary vasculature is its capacity to accommodate blood flow from 6L/ min at rest to 25 L/min under strenuous exercise without changes in pulmonary pressure. The mean pulmonary arterial pressure is 14+3 mmHg.[38] Reduced blood flow and low shear stress (5–8 dyn/cm2 ) suggest pulmonary hypertension. Physiological flow shear stress (10–25 dyn/cm2 ) is an important regulator of gene expression in endothelial cells.[37] Chronic hypoxia can increase smooth muscle in small and non-muscular arteries of the pulmonary circulation. This leads to vascular stiffness and pulmonary hypertension.[38] The pulmonary endothelium presents lipids to type-2 pneumocytes from the blood to produce surfactants.[10,31]
Centrilobular Lung Interstitium
This is loose connective tissue intervening between alveolar pneumocytes and pulmonary capillary endothelial cells. It is functionally a dry tissue space. The lung interstitium comprises type -1 collagen, elastic fibers, and proteoglycans such as versican, perlecan, and syndecan. There is a turnover of > 10% per day of the total pulmonary extracellular matrix.[14]
Interstitial cells include lipofibroblasts, myofibroblasts, pericytes, migratory leucocytes, lymphatics, and mesenchymal stromal cells. These cells form 35% of the alveolar complex.[41] The extracellular matrix is important in maintaining the internal milieu.[14]
It is stronger than the alveolar epithelium and pulmonary endothelium and resists injury till the last. This is because of the components in the form of collagen, laminin, enactin, and heparan sulfate proteoglycans.[41] High altitude and alveolar hypoxia can induce vasoconstriction and extravasation of protein-rich fluid into the interstitium causing pulmonary edema.[31]
The interstitial fibroblast cells
The interstitial fibroblast constitutes 40% of lung cells. It synthesizes the protein and non-protein parts of the extracellular matrix. The fibroblasts are influenced by TGF β 1 and TGF β 3.[14] Computerassisted transmission electron microscopic serial section 3D reconstruction of alveolar wall fibroblasts studies have shown that lung fibroblasts connect pulmonary capillary endothelial cells to type- 2 pneumocytes across the interstitium through apertures of respective basement membranes. This link serves as a conduit for the crawling leucocytes to enter the alveoli of the lungs.[15]
TEM 3D reconstruction of the lung alveoli has revealed two types of fibroblasts. One type lies parallel and another with microfilaments (myofibroblasts) lying perpendicular to the alveolar epithelium. The myofibroblasts bridge the gap between the type-2 pneumocytes and the pulmonary endothelium and pave a continuous pathway for the migration of leukocytes. Disruption of this pathway is responsible for the diseases like emphysema.[15]
Pericytes
Pericytes are mesenchymal cells that resemble smooth muscle cells of a blood vessel. They have cytoplasmic processes that enclose the capillary endothelium. Pericytes adhere to endothelial cells by tight junctions containing fibronectin adhesion molecules and peg and socket association with tight, gap, and adheren junctions.[41] Pericytes and resident fibroblasts help in cellular differentiation during lung development. They can differentiate into osteocytes, chondrocytes, and adipocytes. Hence, pericytes have been indicated in stem cell transplantation and regenerative medicine. They also maintain homeostasis mainly collagen turnover in the lung interstitium.[41] Thus, pericytes maintain vascular integrity and regulate tissue growth patterns.[13]
Pericytes play an integral role in preventing pathological changes in the lung tissue. Decrease in pericyte population causes loss of antibacterial barrier leading to exacerbation of lung pathology. Pericytes induce fibrogenesis as a part of the defense mechanism of the lungs to the infection commonly seen in asthma, COPD etc. Pericytes also play a role in inducing pulmonary hypertension.[42]
Alveolar Macrophages
They belong to the lineage of terminally differentiated myeloid cells. They have long cytoplasmic extensions called pseudopods. They play a crucial role in the phagocytosis of exogenous and endogenous tissue debris.[43]
Alveolar macrophages get activated in response to bacterial infection by producing reactive nitric oxide (NO), destroying the bacteria. In cigarette smokers, there is an increased production of gelatinases (MMP2 and MMP9) and macrophage metalloelastase (MMP12) from alveolar macrophages, the latter destroying lungs.[20]
They can be differentiated from interstitial macrophages by the expression of CD11b and CD68. Once exposed to a pathogen, the cells differentiate into M1 and M2 cells. M1 cells produce TNF-alpha and IL-1beta. M2 cells produce allergic cytokines IL-4, IL-13, and IL 17, involved in the mechanism of bronchial asthma.[41,43]
Summary of morphometric characteristics of alveolar cell complex
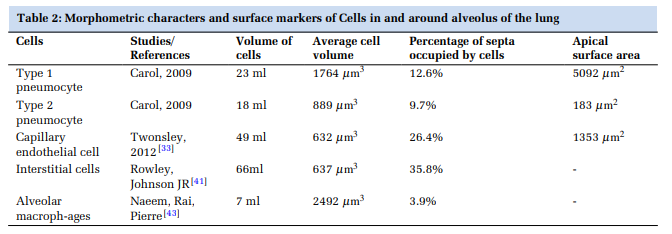
The morphometric characters of the alveolar cell complex have been summarised in Table 2.
Markers of alveolar cell complex and mechanism of injury
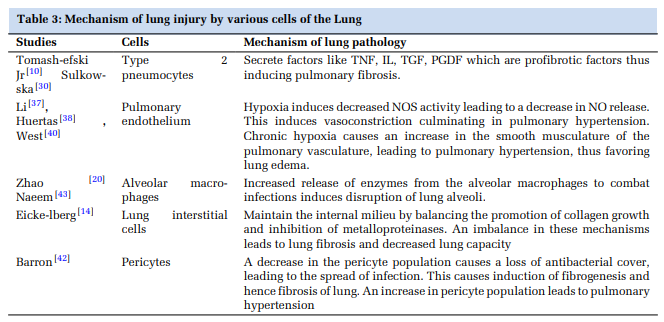
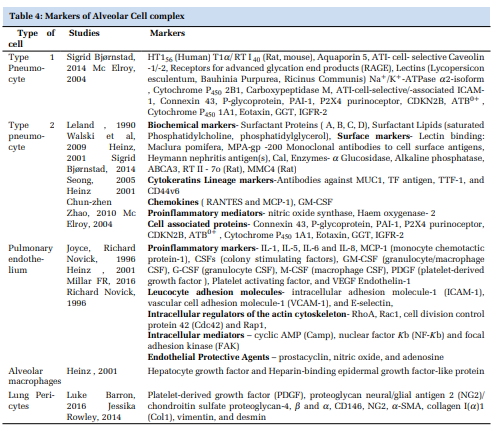
Mechanisms of lung injury caused by various cells of the lung and their markers have been summarized in Table 3 and Table 4, respectively.
The primary function of lung is gaseous exchange facilitated by the type-1 pneumocyte and capillary endothelial combination. The gaseous exchange is enhanced by the elastic expansion of the lung assisted by the surfactant proteins. The type-2 pneumocyte, primarily a surfactant producer, also alters into type-1 pneumocyte as a defense cell influencer.
The interaction between endothelial cells, alveolar macrophages, type-2 pneumocytes, and the interstitium is critical in response to lung injury/infection. The chain of events triggered by the release of chemical mediators by these cells leads to inflammatory edema, respiratory distress, and fibrosis. The extracellular matrix is the strongest and resists injury till the last.
Type 2 pneumocytes are stem cells. Is it the entire population or its subpopulation, which acts like stem cells, the future research area? The role of collateral circulation in interstitial lung diseases needs to be explored. The contribution of the glycocalyx layer of pulmonary vessels in ARDS is still unclear. The fact that type 2 pneumocytes have the ability to interact needs probing. Similarly, the precise role of pulmonary endothelial cells in neutrophil sequestration needs to be explored.
RTI40: rat type I cell 40-kDa protein, TIa: type I cell alpha protein, HTI56: human type I cell 56-kDa protein, Naz/Kz-ATPase: sodium/potassium exchanging adenosine triphosphatase, ICAM1: intercellular adhesion molecule-1, RAGE: receptor for advanced glycation end-products, PAI-1: plasminogen activator inhibitor, CDKN2B: cyclin-dependent kinase 4 inhibitor 2B, ATB0z: Naz- and Cl- -dependent neutral and cationic amino acid transporter B0z, GGT: c-glutamyl transferase, IGFR-2: insulin-like growth factor receptor 2
Subscribe now for latest articles and news.